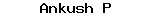
Here is an essay on the ‘Classification of Gymnosperms’ for class 11 and 12. Find paragraphs, long and short essays on the ‘Classification of Gymnosperms’ especially written for school and college students.
Classification of Gymnosperms
Essay Contents:
- Essay on Pteridospermopsida
- Essay on Cycadopsida
- Essay on Ginkgopsida
- Essay on Coniferopsida
- Essay on Gnetopsida
Essay #
1. Pteridospermopsida:
The Carboniferous period is considered as the age of ferns’ because of the presence of a large number of primitive ferns (Primofilicales) as well as fern-like fronds of uncertain affinity. It was Stur (in 1883) who first time hypothesised the existence of such type of plants.
The systematic position of such fern-like plants were not established. Williamson (1887) recognised the existence of such plants showing combined structural characteristics of both cycads and ferns based on anatomical features.
It was H. Potonie in 1899, who, on the basis of anatomical evidence, suggested the existence of a group of plants that was transitional between ferns and seed plants and introduced the concept of Cycadofilicales (Pteridosperms). Finally, two British palaeobotanists, F. W. Oliver and D. H. Scott, in 1904, established the concept of Pteridosperms (seed ferns) based on the detailed work on Lyginopteris oldhamia plant.
The Pteridosperms were a very large and diverse assemblage of plants which first appeared in the Upper Devonian, had its climax during the Carboniferous to Triassic and became extinct in the Cretaceous. Due to their wide span of existence, they are conventionally divided into two broad groups: Palaeozoic Pteridosperms (Upper Devonian to Permian) and Mesozoic Pteridosperms (Triassic to Cretaceous).
General Characteristics of Pteridospermopsida:
1. Some were small trees with upright trunks, others were lianes or vines with scrambling habit.
2. Massive fern-like fronds arranged helically on stem.
3. Primary xylem mesarch (rarely exarch) with variable stelar configurations, ranging from simple protostele to eusteles.
4. Wood is manoxylic and limited in amount. Thin-walled tracheids are with multiseriate pitting on radial walls.
5. Both pollen-bearing organs and ovules (seeds) borne on leaves.
6. Pollen organs aggregated into clusters or arranged into large synangiate organs.
7. Seeds large and solitary, sometimes small and surrounded by cupulate structure.
Order 1: Cycadofilicales (= Pteridospermales)
Family 1. Calamopityaceae:
The family Calamopityaceae is an interesting group of permineralised plants. The family consists entirely of stems and petiole remains where other organs are not known.
The different form-genera so far identified are:
Stems: Stenomyelon, Calamopitys
Petiole: Kalymma
Class. Pteridospermopsida:
Stems:
The Lower Carboniferous stem (Stenomyelon) is considered to be the most primitive member of the family. The beautifully preserved stem (Fig. 1.1 A) of S. tuedianum shows a primitive type of three lobed protostele dissected by the radiating plates of parenchyma like the Aneurophytales of progymnosperm.
Hence this genus is considered to be the ancestor of seed ferns. Metaxylem tracheids exhibit multiseriate bordered pits. In another species, 5. primaevum, the protoxylem is entirely composed of tracheids with mesarch primary xylem.

Calamopitys is the commonest genus showing a large parenchymatous cortex with sparganum cortical sclerenchyma and leaf traces (Fig. 1.1B). The stele appears roughly triangular which ranges from protosteles with abundant parenchyma to eusteles. C. americana is a common species that exceeds 4.0 cm in diameter. The pith is of mixed type, comprised of tracheids and parenchyma.
In both the genera, stems are with compact manoxylic wood, surrounded by primary xylem.
Petiole:
The petioles (Kalymma) are attached in 2/5 phyllotaxy to the stem of Stenomyelon, showing many traces containing two exarch primary xylem. Some petioles show a number of bundles that assume a C-configuration.
Family 2. Lyginopteridaceae:
The family Lyginopteridaceae shows its historical importance in the recognition of the group Pteridosperm (seed fern).
There are some characteristic features which can be used to distinguish the family from rest of the members of Pteridosperms:
1. Presence of cauline monostelic vasculature.
2. Presence of axillary branching.
3. Fronds are large with bifurcate rachis.
4. Petioles are small with a single V-, Y- or W-shaped trace those are formed from the fusion of several small traces.
5. Pollen-bearing organs are small, laminar, or terminal in clusteristics on branches of fronds.
6. Pollens are of pre-pollen type with trilete aperture.
7. Ovules are small, radially symmetrical (radiospermic) and borne in cupules having hydrasperman pollen chamber.
8. Nucellus is fused with integument and apex forming a salpinx or lagenostome.
The several important form-genera of Lyginopteridaceae so far identified are:
Stems: Lyginopteris, Heterangium, Microspermopteris, Schopfiastrum, Tristichia.
Frond: Sphenopteris.
Root: Kaloxylon.
Rachis: Rachiopteris, Lyginorachis.
Pollen-bearing organs: Crossotheca, Telangium, Telangiopsis, Feraxotheca.
Seeds: Lagenostoma, Physostoma, Stamnostoma, Eurystoma, Lyrasperma, Sphaeros-toma, Conostoma.
Cupule: Calymmatotheca.
Family 3. Medullosaceae:
The members of this family are known to occur in the Mississippian (Lower Carboniferous) extending up to Permian. They were more massive than the Lyginopteridaceae. The genus Medullosa was initially established by Cotta in 1832 for Lower Permian stem fossils.
The distinguishing characteristic features of the family:
1. Plants are massive trees.
2. Stems with several segments of vascular tissue, each surrounded by secondary xylem.
3. Fronds with a bifurcate rachis.
4. Massive petioles with large number of scattered traces.
5. Pollen organs synangiate with tubular sporangia.
6. Pollens are of pre-pollen type, large with monolete aperture.
7. Seeds are large, non-cupulate, three- angled.
8. Nucellus free from integument, except at the base.
9. Pollen chambers simple.
The different form-genera of this family so far identified are:
Stems: Medullosa, Colpoxylon, Sutcliffia, Quaestora.
Leaves: Neuropteris, Alethopteris, Reticulopteris, Callipteridium, Mixoneura, Linopteris, Cyclopteris, etc.
Petiole: Myeloxylon.
Pollen organs: Codonotheca, Whittleseya, Aulacotheca, Dolerotheca, Bernaultia, Potonie, etc.
Ovules (seeds): Pachytesta (= Trigonocarpus), Stephanospermum, Aetheotesta, Codonospermum, etc.
Family 4. Callistophytaceae:
The family Callistophytaceae is probably the best-known group of Palaeozoic Pteridosperms. Callistophytean fossils are principally known from Middle to Upper Pennsylvannia of USA and Upper Carboniferous of France.
General characteristic features of Callistophytaceae:
1. Small shrubby, scrambling plants that branch profusely.
2. Stem eustelic that bears spirally arranged pinnately compound leaves with axillary buds or branches at each node.
3. Young leaves show circinate vernation.
4. Pollen organs synangiate and borne superficially on pinnules of the frond.
5. Pollen grains are small and monosaccate.
6. The seeds are small, platyspermic and non-cupulate.
7. The integument is free from the nucellus except at the base.
The different form genera of this family:
Stems: Callistophyton
Frond: Medullopteris.
Pollen organs: Idanothekion, Callandrium
Dispersed pollen: Vesicaspora.
Seeds (ovules): Callospermarion.
Order 2: Glossopteridales:
Family: Glossopteridaceae:
The name Glossopteris was first proposed by Brongniart (1828) for certain tongue-shaped leaves with a prominent midrib and reticulates venation. Glossopteris occurs in Australia, Africa, South America, Antarctica and India and constitute the Permian Condwana land floras of Southern Hemisphere that separated from other continents of Northern Hemisphere by the great Tethys sea.
Glossopteris first appears in the Upper Carboniferous, forms a climax vegetation during the Permian and decline thereafter. Some specimens are also known from the Triassic period of India and the Jurassic deposits of Mexico.
From the evidences obtained from the works of various workers, the Glossopteridales has been characterised by the following features:
1. The Glossopteris was a large tree (Fig. 1,9A). The trunk was about 6 m tall with araucarioxylon type of gymnospermous wood. The large number of isolated leaves and, to a much lesser extent, reproductive organs, suggest that the plants were deciduous. This has been established by the occurrence of numerous isolated leaves that are found in autumn/winter sediments, whereas they are absent in spring/summer sediments.
2. The plant was supported by a root system of Vertebraria type.
3. The secondary growth was present in stems, branches and roots showing conspicuous growth ring that indicates climatic variations.
4. Leaves were arranged alternately or in tight helices to form whorls on short shoots.
5. Reproductive organs were either borne on leaves or attached to highly modified leaves. Both male and female fructifications were borne separately. It is not known whether they were monoecious or dioecious?

Different important form-genera of Glossopteris plant so far identified are:
1. Root: Vertebraria
2. Leaves: Glossopteris, Gangamopteris, Palaeo- vittaria, Rubidgea, Euryphyllum, Rhabdotaenia, etc.
3. Male fructifications: Eretmonia, Glossotheca, Kendostrobus.
4. Ovule structures: Lidgettonia, Ottokaria, Scutum, Denkania, Dictyopteridium.
Order 3: Caytoniales:
Caytoniales is another group of Pteridosperms which was widespread during the Upper Triassic to Lower Cretaceous. Initially Caytoniales was considered as a close associate of angiosperms because of the apparent closed carpel-like cupulate structure. But now the members of Caytoniales are included under Pteridosperms and are divided into three distinct families namely, Caytoniaceae, Peltaspermaceae and Corystospermaceae.
Family 1. Caytoniaceae:
The family Caytoniaceae had a wide strati- graphic range (U. Triassic to L. Cretaceous). The leaves of this family are placed in form-genus Sagenopteris, the male fructification in Caytonanthus and the female fructification in Caytonia. These form genera were found in close association in the deposits of Greenland, Western Canada, Eastern Russia, England, Siberia, etc.; however, no organic connection could be noticed.
i. Leaf:
Sagenopteris leaf (Fig. 1.10 A) is palmately compound, consisting of two pairs of four lanceolate leaflets, with a slender petiole. Each leaflet has a prominent midrib showing reticulate venation. Haplocheilic stomata are found on the lower surface.
ii. Male Fructification:
The male fructification, Caytonanthus (Fig. 1.10 B), is a pinnately branched structure which bears synangia (four fused sporangium) at its ultimate division. A single synangium is 1.0 cm long, radially symmetrical, pendent, tubular and quadrilocular (Fig. 1.10C). Pollen grains are small and bisaccate with a distal sulcus aperture, referred to the form-genus Vitreisporites. The sporoderm (exine) is bilayered, the inner nexine lamellate and the outer sexine alveolate.
iii. Female Fructification:
Caytonia (Fig. 1.10D) is a flat structure which bears fruit, rachis or megasporophyll. The rachis is considered as megasporophyll, because of its dorsiventral epidermal structure. The seeds are completely enclosed within the fruit-like body or cupule with a minute opening or crack below, close to the pedical (Fig. 1.10 E).
There are a series of transversely arranged thickened bars on the curved inner face of a well- developed cupule (Fig. 1.10 F), and the number of these bars corresponds with the number of seeds. Generally 8 to 30 orthotropous seeds are present in single or double rows.
Each seed is flattened, oval and unitegmic (with one integument) with a short micropyle pointing towards the opening below. The nucellus is free from the integument. The integument is devoid of any vascular trace.
Caytonia was compared with the angiospermc by the earlier workers due to the presence of fruit-like structure, the reticulate venation of leaves and the microsporophyll bearing tetralocular sporangia. Hence, Caytoniacene has been considered as the probable ancestor of angiosperms.
However, Caytoniaceae exhibit many non-angiospermous characteristics:
(a) Radially symmetrical pollen bearing organ and lack of a filament and connective,
(b) No blind vein ends in leaf,
(c) Bisaccate pollen grains with alveolate sporoderm,
(d) Flate, pinnate megasporophyll,
(e) Single integument in ovule,
(f) Pollen reached the micropyle by means of a “pollen drop mechanism” as in gymnosperms.
The Caytoniaceae has now been considered as an unique member of gymnosperms and treated under a separate and independent order Caytoniales of Pteridospermopsida.
Family 2. Peltaspermaceae:
The Peltasperms are a small group of plants known from Upper Triassic deposits of South Africa, Greenland and Sweden. The genus Lepidopteris has been designated for the whole plant in spite of having several form genera, because all the form genera have the characteristic blister-like swelling and similar stomatal structures.
The different form-genera so far identified are:
i. Leaf: Lepidopteris
ii. Male fructification: Antvesia
iii. Female fructification: Peltaspermum.
i. Leaf:
The frond of Lepidopteris are bipinnate with broadly based, open-veined pinnules. The rachis and pinnules of the fronds are provided with blister-like swelling.
ii. Male Fructification:
The pollen organs, Antvesia, are pinnately branched which give two to three dichotomously dividing ultimate branches. Each ultimate branch bears two rows of pendulous pollen sacs. Pollen grains are non-saccate, bilaterally symmetrical with a longitudinal furrow.
iii. Female Fructification:
The ovuliferous structures are very distinctive, called Peltaspermum. About 20 ovules (seeds) are borne in a ring on the underside of a stalked, peltate disc. Several discs are helically arranged on a central stalk. Each ovule (seed) is 7 mm long with an elongated and curved micropylar beak.
Family 3. Corystospermaceae:
The leaf genera, Dicroidium of Corystospermaceae was first described by H. H. Thomas (1933) from Triassic beds of Natal, South Africa. Most of them were small plants with pinnatified and dichotomously veined leaves and only known from Gondwanaland.
The principal and best-known form genera so far reported are:
i. Leaves: Dicroidium (= Thinnfeldia), Pachypteris. Stenopteris.
ii. Male fructification: Pteruchus.
iii. Female fructification: Umkomasia
Although no parts of the plants have ever been found attached, they are believed to belong to Corystospermaceae on the evidence of similar epidermal characteristics.
i. Leaves:
All the three leaf genera, (Dicroidium, Pachypteris, Stenopteris) show pinnate frond with open venations (Fig. 1.101 A, B). Pigg (1988) found organic connection between Dicroidium and ovulate cupules from Triassic deposits of Antarctica. Internal structure is similar to that of cycads. A single rachis bears 15 to 20 bundles arranged in a ring of 7 to 8 near abaxial surface and a line of 5 to 8 adaxially. Mesophyll is differentiated into palisade and spongy parenchyma. The epidermis is provided with dicyclic stomata.

ii. Male Fructification:
The pollen-bearing organ, Pteruchus is a small branching system, divided by equal and unequal dichotomies. Each microsporophyll terminates in a flattened, rounded head which bears clusters of elongated pollen sacs on its underside. Pollen grains are bisaccate having a well-defined aperture.
iii. Female Fructification:
The megasporophyll, Umkomasia is situated in the axil of a bract. Each megasporophyll is branched and is subtended by a pair of bracteole. Each branch bears opposite pairs of stalked, recurved, bivalved, helmet-shaped (Corystosperm means helmet-shaped seed) cupule. Each cupule bears one seed. The micropyle is curved and bifid, projecting beyond the cupule.
Essay #
2. Cycadopsida:
The class Cycadopsida includes two orders : Cycadales and Cycadeoidales. They were the most dominating taxa during mid-Mesozoic Era. However, they were first appeared in the Upper Carboniferous period of Palaeozoic Era and declined in the late Cretaceous.
General Characteristics of Cycadopsida:
1. Plants are generally with\stout trunk having manoxylic wood.
2. Leaves are large frond-like, they are basically pinnate in form or in venation.
3. Seeds are large showing radial symmetry.
4. Presence of ciliated sperms.
Order 1: Cycadales:
The members of the Cycadales are woody plants with unbranched stem (occasional with adventitious branching). They bear manoxylic wood with mucilage canals in pith and cortex. Leaves are large, pinnate (rarely bipinnate) with haplocheilic stomala. Leaf trace is of diploxylic nature (except in Nilssoniaceae). Plants are dioecious and reproductive organs in cones (except female Cycas). Megasporophylls bear 2-8 ortho- tropous ovules with sterile tip. Seeds are large. Microsporophylls are scale-like or peltate with abaxial sporangia. Sperms are multiciliated.
Most authors recognised two families in the order — Nilssoniaceae and Cycadaceae. However, some authors classified the order into four families, such as Nilssoniaceae Cycadaceae, Stangeriaceae and Zamiaceae. Some others preferred to retain a single family Cycadaceae.
The members of Cycadales are commonly called as ‘cycads’ and are the only living representatives of the class. The Cycadales probably originated from the Pteridosperm ancestor (Palaeocycas megasporophyll with Taeniopteris leaf) in the Upper Carboniferous to Permian period. They reached their climax during the Triassic to Jurassic period of Mesozoic Era and mostly declined at the end of Upper Cretaceous period. Thus, the Mesozoic Era is commonly known as “age of cycads”. At present, they are represented by eleven extant genera and are restricted in the tropical zones of both the Eastern and Western Hemispheres.
The earliest known cycads have been reported mostly as dispersed megasporophylls. Spermopteris (Fig. 1.11 A) is the first evidence of the fossil cycad from the Upper Pennsylvania of Kansas. It is an ovulate structure of the leaf genus Taeniopteris where two rows of ovules attach to the abaxial surface of the leaf.
Plasmatocycas and Archaeocycas are the two cycadean megasporophylls reported from the Lowen Permian. Plasmatocycas (Fig. 1.11B) megasporophyll has two rows of ovules attached laterally to the stalk of a Taeniopteris leaf. Archaeocycas (Fig. 1.11C) megasporophyll has an expanded spathulate lamina bearing two rows of ovules on the basal part. The ovules are partly enclosed by the lamina in the basal region.
Leptocycas gracilis is an Upper Triassic cycad, reconstructed by Delevoryas and Hope (1971). The plant has a smooth, slender stem (1.5 m tall) bearing a crown of fronds and a pollen cone.
The reconstruction of Bjuvia simplex (Fig. 1,11D), an Upper Triassic cycad, has been made by Florin (1933) based on the Taeniopteris leaves and Palaeocycas megasprophylls with an imaginary trunk.
Harris (1961) reconstructed a Jurassic cycad bearing Beania gracilis ovulate cones (Fig. 1.11E, F) and crown of Nilssonia type (Fig. 1.1.1 G) leaves on an imaginary stem. The ovulate cones of Beania gracilis resemble seed cone of Zamia. The pollen cone, Androstrobus (Fig. 1.11 H, I) is believed to be associated with the same plant.
The cycads comprise of eleven (11) living genera of which five (Microcycas, Dioon, Chigua, Ceratozamia and Zamia) belong to Western Hemisphere and the remaining six (Cycas, Stangeria, Bowenia\ Macrozamia, Lepidozamia and Encephalartos) are confined to Eastern Himesphere. A total of about 100 species of cycads have been reported.

Order 2: Cycadeoidales (= Bennettitales):
The Cycadeoidales (= Bennettitales) is an extinct group of fossil gymnosperms. The order was named Bennettitales in honour of the English botanist J. J. Bennett. This order caught attention during earlier times by the discovery of silicified trunk known as Cycadeoidea from Great Britain.
At the same time, Bennettites plant was reported from the Lower Cretaceous deposits of Isle of Wight. It becomes apparent that there is no difference between Cycadeoidea and Bennettites. Thus, according to the Rule of Priority Cycadeoidea is a valid name for the genus.
Both the members of Cycadales and Cycadeoidales coexisted during Mesozoic Era. The Mesozoic Era is thus popularly called the “age of the cycads.” Cycadeoid leaves are very much similar to cycads in form, structure and venation. However, cycadeoid leaves differ from those of cycads in having syndetocheilic stomata, while it is haplocheilic in cycads. Moreover, cycadeoid guard cells are heavily cutinised than that of cycads.
General Characteristics of Cycadeoidales:
1. In some members, plants are with slender and highly branched stems, or others have trunk-like stems with few branches, or are unbranched.
2. The stem surfaces are covered with spirally arranged persistent leaf bases with or without hairs in-between.
3. The stem anatomy shows a large pith surrounded by a cylinder of wood.
4. The wood is well-developed and compact like that of conifer wood.
5. The primary xylem is endarch. The secondary tracheids either have scalariform pitting on the walls, or have multiseriate bordered pits on their radial wall.
6. Secretory canals are present in pith and cortex.
7. The leaves are pinnately compound (rarely, simple) with paralled venation and syndetocheilic stomata. The guard cells are heavily cutinised and the stomata are oriented at right angles to the veins.
8. The leave traces originate from the primary xylem and pass directly through the secondary xylem to the cortex where they branch forming many traces. Thus, a single leaf base receives many vascular traces.
9. The reproductive structures are bisporangiate (except, Williamsoniaceae where they are monosporangiate) — often called flowers, protected by many bracts.
10. The ovules are borne in large numbers on a dome-shaped, cylindrical or conical receptacle.
11. The ovules are interspersed with vascularized interseminal scales with their enlarged tips fused to form a shield through which micropyles come out.
12. Each ovule has a single integument which is three-layered and the integument protects forward to form a long micropyle.
13. Microsporophylls are in whorls or free or united with numerous elongated micro- sporangia usually fused into a synangium.
14. The presence of monocolpate (= monosulcate) pollen grains.
15. The embryo is dicotyledonous.
Classification of Cycadeoidales:
Sporne (1965) classified the order Cycadeoidales into three families — Williamsoniaceae, Wielandiellaceae and Cycadeoidaceae.
However, in recent times, the Cycadeoidales has been divided into two families — Williamsoniaceae and Cycadeoidaceae.
The order is comprised of four reconstructed members — Williamsonia, Williamsoniella, Wielandiella and Cycadeoidea.
Family 1: Williamsoniaceae:
Several plants under this family were reconstructed. Williamsonia gigas was reported by Williamson (1870) for the first time from Jurassic of Yorkshire. The most important plant, Williamsonia sewardiana was reconstructed by Prof. Birbal Sahni (1932) from the Jurassic (Upper Gondwana) deposits of the Rajmahal Hills of India.
The oldest known member of this family is Wielandiella which was recovered from the Upper Triassic (Rhaetian) of Scania in Sweden by Nathorst (1911). Another member, Williamsoniella was reported by Thomas (1915) from the Jurassic beds of Yorkshire.
a. Wielandiella:
The plant consists of a very slender, herbaceous stem showing false dichotomy (Fig. 1.25). The unipinnately compound leaves are known as Anomozamites which are concentrated in the region of bifurcations.
A single sessile bisporangiate cone is borne in the region of each bifurcation. The compression-impression cone shows a central ovulate receptacle bearing ovules, interseminal scales and microsporophylls. The cone is subtended by numerous bracts.


b. Williamsoniella:
It is a shrubby plant that bears dichotomously branched axes (Fig. 1.26). The simple, petiolate and lanceolate leaves under the form-genus Nilssoniopteris are borne scattered on the axes.
Each bisporangiate cone is borne in the apex of peduncle that develops in the axil of a leaf. There is a centrally elongated ovulate receptacle that bears about 300 ovules and interseminal scales. In the basal region of the receptacle, there are 12 or more microsporophylls and numerous bracts arranged in whorl.
c. Williamsonia:
Williamsonia is the widely studied genus of the family Williamsoniaceae. Several species of Williamsonia have been described from different regions of the world.
Birbal Sahni (1932) reconstructed Williamsonia sewardiana (Fig. 1.30) using Bucklandia indica (stem), Ptilophyllum cutchense (leaf) and Williamsonia (female fructification). T. M. Harris (1969) also made a reconstruction using Bucklandia pustulosa (stem), Ptilophyllum pecten (leaf) and Williamsonia leckenbyi (female fructification).
The male fructification of Williamsonia is called Weltrichia which has never been found in actual connection with the plant. Several species of Weltrichia are described from the Jurassic deposits of India. In addition to Ptilophyllum, several leaf-genera have also been described showing cycadeoid affinity.
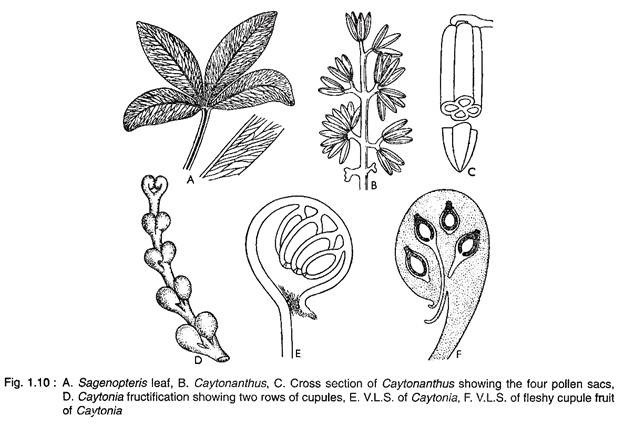
Leaves:
Leaves are pinnately compound. Many long slender pinnae are attached to the adaxial surface of the rachis by full width of their broad bases. Pterophyllum (Fig. 1.27A, B) leaf is characterised by long, slender pinnae with paralled venation, attached laterally to the rachis. Ptilophyllum (Fig. 1.27C, D) leaf has linear to slightly sickle-shaped pinnae, with parallel venation attached to the adaxial surface of the rachis.
Otozamites pinnae (Fig. 1.27E, F) have lobed bases with spreading veins which are attached to the adaxial side of the rachis. Dictyozamites leaf is similar to Otozamites except that the pinnae have anastomosing network.
Stem:
The stem, called Bucklandia is columnar in nature, either unbranched or sparsely branched. The surface of the stem is covered with spirally arranged persistent leaf bases. Internal structures are rarely preserved, because most of the specimens were recovered as casts. Anatomically, the stem exhibits a broad cortex, vascular bundles arranged in a ring and a narrow pith. The wood is formed following secondary growth in thickness.
Male Fructification:
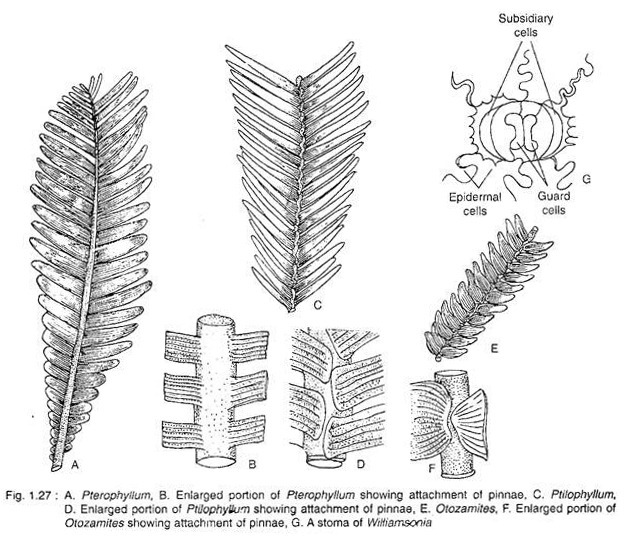
The male fructification called Weltrichia have never been found in association with Williamsonia. However, their attachment with Williamsonia has been inferred by the association of a portion of other form genera and also by having syndetocheilic stomata (Fig. 1.27C).
Several microsporophylls are arranged in a whorl and fused at the base forming cup-shaped (Weltrichia whitbiensis) (Fig. 1.28C) or bell- shaped (W. spectabiiis) (Fig. 1.28A) structure. Each microsporophyll has two rows of synangium bearing branches on the inner surface (Fig. 1.28B). The pollen grains that produced within sporangia are of monocolpate (= mono- sulcate) type.
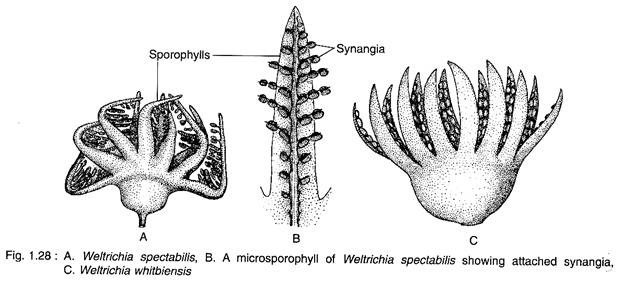
Female Fructification:
Several species of Williamsonia have been described. The female fructification consists of a well-developed central conical receptacle which bears numerous stalked ovules and interseminal scales arranged spirally on the receptacle (Fig. 1.29). The interseminal scales are vascularised, club-shaped fleshy structures.
The distal club-like ends of interseminal scales are fused to form a shield through which ovular micropyles protrude. The female fructification is subtended by spirally arranged perianth-like bracts with scales at their base and hairs above.

d. Williamsonia Sewardiana:
One of the most important contributions made in Palaeobotany is the reconstruction of Williamsonia sewardiana by Prof. Birbal Sahni (1932) from the Jurassic (Upper Gondwana) deposits of the Rajmahal Hills of India.
The plant was named in the honour of two English botanists Prof. Williamson (grand-teacher of Dr. Sahni) and Prof. A. C. Seward (teacher of Dr. Sahni). It was the first reconstructed fossil plant from India. The name of this plant is actually based on the female fructification (Williamsonia) rather than the entire plant of which Bucklandia indica is the stem and Ptilophyllum cutchense, the leaves.
The reconstruction as made by Dr. Sahni shows a small tree-like plant with sparsely branched columnar stem (Bucklandia indica) ending in a crown of pinnately compound leaves (Ptilophyllum cutchense) and bearing female fructification (Williamsonia scotica) on short branched lateral shoot. The plant looks like a miniature Cycas (Fig. 1.30).

e. Bucklandia Indica:
The stem is armoured with spirally arranged alternating areas of large (foliage) and small (scaly) leaf bases (Fig. 1.30). The branching of the stem is lateral which projects beyond the armour of leaf-curshions. There are two types of branching the vegetative or sterile shoot and the fertile shoot.
There is a prominent constriction at the base of each lateral shoot. It is presumed that the lateral shoots get detached through the constricted base and vegetatively propagate. The main shoot bears both the foliage leaves and scale leaves, while the lateral shoots bear only foliage leaves.
Internally, the stem shows a wide cortex and large pith. The vascular bundles are conjoint, collateral, endarch and open, remain embedded in pith. Many secretory ducts are found in cortex and pith. The secondary xylem, although compact, is manoxylic comprising of scalariform tracheids with multiseriate bordered pits on radial wall.
There are uni- to triseriate medullary rays. There are numerous leaf traces. The most striking difference with the Cycas stem is the absence of girdling leaf traces in Bucklandia indica.
f. Ptilophyllum Cutchense:
The fronds have linear to slightly sickle- shaped pinnae which are attached to the adaxial surface of the rachis by full width of their broad and rounded bases (Fig. 1.27C, D). Pinnae exhibit parallel venations.
Leaf traces arise from the primary xylem and pass directly through the secondary xylem to the cortex where they again branch to form many traces. Thus, a single leaf base receives many leaf traces. The leaves exhibit syndetocheilic stomata with heavily cultinised guard cells. The stomate are oriented at right angles to the veins. The epidermal cell walls have wavy outlines (Fig. 1.27G).
g. Williamsonia:
Based on the similar anatomical features in the peduncle of Williansonia scotica cone and Bucklandia indica stem, Dr. Sahni was able to deduce a connection between these two form- genera. Thus, the flower that attached to the reconstructed plant Williamsonia sewardiana is supposed to be Williamsonia, a female fructification.
Williamsonia consists of a well-developed central conical receptacle bearing numerous spirally arranged stalked ovules and interseminal scales. The interseminal scales are much more in number than that of ovules (Fig. 1.29). The tip of the receptacle remains open. The ovules are orthotropous and shortly stalked. The nucellus is fused with the single integument except at the top.
The integument extends to form a long and narrow micropylar canal. The interseminal scales are vascularised, club-shaped fleshy structures. Their distal club-like ends are provided with stomata and are fused to form a shield through which ovular micropyles protrude. The entire flower is sublanded by spirally arranged bracts with scales at their base and hairs above.
Family 2: Cycadeoidaceae:
Cycadeoidea is the only genus under the family Cycadeoidaceae and has a wide geographical distribution, especially in North America, Europe and India during Upper Jurassic to Upper Cretaceous. The plants have stout, globose trunks with armour of spirally arranged leaf bases embedded in ramenta of flat, tongue- shaped scales (Fig. 1.31). The leaves have never been found attached to the stem.
It is assumed that Ptilophyllum or Dictyozamitestype of leaves were borne spirally in a crown at the top of the trunk. The flowers were borne in the axil of leaf bases and developed simultaneously. Thus the plants were monocarpic i.e., they flowered once during their lifetime.

Anatomically, the trunk exhibits a large pith surrounded by a broad ring of vascular tissue comprising of manoxylic wood and secondary phloem (Fig. 1.32). The vascular ring is dissected by broad rays, either uniseriate or biseriate type. There are scattered secretory ducts in cortex and pith.
The leaf traces traversing the cortex are C- shaped and pass directly through the secondary xylem to the cortex where they again divide and arrange themselves in a horse-shoe-shaped fashion without girdling.

Cycadeoid flowers are bisexual, borne on a short pedical and are covered by about 100 spirally arranged hairy bracts, protected by a ramentum or scales (Fig. 1.33A). The bracts open up at maturity. There are about 20 pinnate microsporophylls arranged in a whorl at the base of ovuliferous receptacle. Microsporophylls are united at the base and comparable to the segments of an orange.
Two rows of kidney- shaped synangia are borne on the inner surface of each pinnule of a microsporophyll (Fig. 1.33B). Each synangium bears 20-30 tubular pollen sacs or sporangia containing monocolpate (monosulcate) pollen grains.
As in Williamsonia, the ovule- bearing structure is in the form of a conical receptacle on which ovules and interseminal scales are arranged spirally (Fig. 1.33C). The number of ovules is almost equal to the number of interseminal scales.

Essay #
3. Ginkgopsida:
The class Ginkgopsida includes a single order Ginkgoales – now represented by a single monotypic extant plant, Ginkgo biloba. The members of this group first appeared in the Permian period. They were well represented all over the world during the Jurassic period and gradually started declining from Cretaceous onwards. The extant plant, Ginkgo biloba, has its fossil record as far back as Triassic.
The leaf impressions of Ginkgo biloba have been identified from the Permian rocks and probably from the Carboniferous rocks. Ginkgo biloba has escaped extinction and still exists today being the oldest living seed plant. Thus, Ginkgo biloba is referred to as ‘living fossil’.
General Charcteristics of Ginkgopsida:
1. Mostly tall woody branching trees.
2. Secondary wood pycnoxylic.
3. Leaves are leathery, strap-shaped or fan-shaped, often deeply divided into dichotomous venation.
4. Male frutifications are axillary, unbranched, catkin-like bearing microsporangiophores.
5. Each microsporangiophore bears 2-12 pendulous microsporangia.
6. Presence of ciliated sperms.
7. Ovules 2-10, terminal on axillary branching or almost unbranched axes.
8. Seeds are large showing radial symmetry.
9. Seeds are with fleshly outer layer and stony middle layer.
Order: Ginkgoales:
The order Ginkgoales is comprised of two families – (i) Trichopityaceae, (ii) Ginkgoaceae.
The family Trichopitys consists of a single genus Trichopitys and is probably the oldest known Ginkgophytes, reported as far back as Permian. The family Ginkgoaceae is represented by 14 extinct genera (e.g. Baiera, Arcrobaiera, Sphenobaiera, Ginkgoites etc.) are by one living genus, Ginkgo. The extinct members (Fig 1.34 A,B) showed their worldwide distribution during Jurassic.
Essay #
4. Coniferopsida:
The Coniferopsida includes four orders:
Cordaitales, Voltziales, Coniferales and Taxales. They probably emerged during the Upper Devoniian and were the most dominating taxa during Upper Carboniferous to Triassic. At present, this class is represented by 57 living genera.
General Characteristics of Coniferopsida:
1. Mostly tall woody plants which are profusely branched.
2. Secondary wood pycnoxylic.
3. There are needle-shaped, paddle-shaped or fan-shaped leaves which are basically dichotomous in form or in venation.
4. Cones compound unisexual.
5. Seeds bilaterally symmetrical.
Order 1: Cordaitales:
The Cordaitales, a group of extinct plants, is reported to be the world’s first great forest-maker. This group originated during Upper Devonian and formed the vast forests in the Upper Carboniferous and declined at the beginning of Triassic.
The various form genera of Cordaitales have been reported from China, Siberia, North America, Europe, Australia and India. Indian Cordaitales are represented by impression-compression of leaves, seeds and permineralised woods.
Previously, the order Cordaitales was classified into three families — Cordaitaceae, Poroxylaceae, and Pityaceae. Both Poroxylaceae and Pityaceae show their close affinity to pteridosperms. Now, the family Poroxylaceae has been included under the order Pteridospermales and the family Pityaceae shows its close association with pteridosperms. Thus, the order Cordaitales comprises of a single family Cordaitaceae.
General Characteristics of Cordaitales:
1. The plants are tall trees (Fig. 1.50) with slender branched monopodial stem, attaining a height of about 30 m and bear a dense cluster of branches at the top.
2. The roots are highly branched and swallow, forming stilt roots like those of mangroves.
3. The leaves are simple, sessile, linear or lanceolate or spathulate and are spirally arranged.
4. The leaves are without midribs and with parallel venation. There are double leaf traces.
5. The primary vasculature is of siphonostelic forming a narrow band between the large pith and secondary wood.
6. The secondary wood is pycnoxylic.
7. The presence of discoid pith which shows large air chambers separated by diaphragms.
8. The cones are unisexual and compound consisting of a main (primary) axis bearing several spirally arranged secondary shoots subtended by bracts.
9. Each secondary shoot is fertile, consisting of spirally arranged sterile and fertile scales.
10. There is a homology between male and female cones, the female secondary shoot comprises of 1 -4 fertile scales that terminate to 1-4 ovules, and the male secondary shoot with 4-6 terminal pollen sacs.
11. The pollen grains are monosaccate (with one wing).
12. The seeds are bilateral i.e., platyspermic.

Different form genera of this family:
1. Root: Amyelon
2. Stem: Mesoxylon, Pennsylvanioxylon
3. Leaf: Cordaites
4. Fructifications: Cordaianthus
(Both male and female)
5. Seed: Cardiocarpus, Mitrospermum
The main plant is called Cordaites, based on the nomenclature of its leaf genus.
Order 2: Coniferales:
The members of the Coniferales are commonly called conifers. They are the largest order among gymnosperms, represented by 52 genera and about 600 species.
General Characteristics of Conifers:
1. They are woody plants showing profused branching often with long and dwarf shoots.
2. The leaves are dimosphic having both the scale leaves and foliar leaves. Foliar leaves are needle-shaped, paddle-shaped or fan- shaped.
3. The tap root system is often associated with mycorrhiza, thus the roots are called mycorrhizal roots.
4. Secondary wood is pycnoxylic, made up of tracheids with large uniseriate or rarely multi- seriate pits on the radial walls and small wood rays. Fibres are absent.
5. Reproductive structures are unisexual compact cones.
6. Female cones are fundamentally compound, consisting of a main cone axis with a few to many spirally arranged bract scales, each subtending or fused with one ovuliferous scale. Each ovuliferous scale bears two or more (rarely one) ovules on adaxial surface.
7. Male cones are simple, consisting of a cone axis on which few to many scale like micro- sporophylls are spirally arranged. Each microsporophyll bears two to many fused or free pollen sacs.
8. Pollen grains are winged or non-winged.
9. Embryos are with two to many cotyledons.
The order coniferales is divided into six families:

Among the 52 genera, about 30 genera are confined to the Northern Hemisphere and 14 genera to the Southern Hemisphere. The rest of the genera are distributed in both the Hemispheres.
At present, the conifers form the world’s most extensive temperate evergreen forests with exception of a few deciduous taxa like Larix, Taxodium, Metasequoia and Pseudo- larix. The oldest living plants on our planet are found among conifers.
A tree of Pinus longaeva growing in Eastern Nevada, USA is about 4,900 years old and a tree of Pinus aristata in Inyo National Forests of California, USA, has been determined to be more than 4,600 years old.
The giant sequoias like Sequoia sempervirens and S. wellingtonia in the coastal forests of Western North America have been estimated to be over 3,000 years old. A gaint cypress, Taxodium mucronatum, near Oaxaca, Mexico has been determined to be more than 2,000 years old.
The giant sequoias (Sequoia sempervirens and S. wellingtonia) are regarded as the tallest living trees (redwood trees) in the world. Some trees in Sequoia National Park, California have been reported to be more than 80 m in height with a base circumference of 285 m.
The conifers have evolved from the Permo- Carboniferous Voltziales members and they reached their climax in mid-Mesozoic forming extensive forests in Northern Europe.
Among the living families, Pinaceae and Araucariaceae are the more primitive which have evolved probably in Triassic age; while Cupressaceae and Cephalotaxaceae are comparatively younger, probably appeared during Upper Jurassic to Lower Cretaceous period.
Essay # 5. Gnetopsida:
This class Gnetopsida is considered to be the highly evolved group of gymnosperms. The class comprises of three distinct orders: Ephedrales, Welwitschiales and Gnetales with monogeneric family in each. However, some scientists referred to retain them in single order Gnetales with a monogeneric family in each.
General Characteristics of the Class Gnetopsida:
1. Opposite or whorled leaves.
2. Vessels in the secondary xylem.
3. Absence of resin canals.
4. Compound male and female strobili resembling inflorescence.
5. Male flowers with perianth.
6. The microphyle projecting as long tube by prolongation of inner integument.
7. Embryo with two cotyledons.
Order: Gnetales:
The order Gnetales includes trees or lianes, distributed in the tropical and humid regions of the world. The leaves are large, dorsiventral with unicostate reticulate venation. The shoot apex exhibits a definable tunicea-corpus layers. The female gametophyte is tetrasporic. Archegonia are completely absent. There is no free-nuclear phase in the embryogenesis.
The order Gnetales does not show any fossil counterparts, hence it is considered as a recently evolved group of gymnosperms.